

1. INTRODUCTION
Eleutherine bulbosa (Mill.) Urb is a bulbous herb belonging to the Iridaceae family, rich in secondary compounds such as phenolic and flavonoid derivatives, naphthalene, anthraqui-none, and naphthoquinone (Sun et al., 2024). As a traditional remedy, the bulb has been used to treat a number of conditions, such as diabetes, breast cancer, high blood pressure, and sexual issues (Kamarudin et al., 2020; Naspiah et al., 2014). Numerous investigations have confirmed the pharmacological applications of E. bulbosa, particularly its antibacterial, antihypertensive, antidiabetic, anti-inflammatory, and antiviral properties (Da Silva et al., 2024; Harlita et al., 2018; Herman et al., 2024; Jiang et al., 2020; Panyachariwat et al., 2024). Given its medicinal and other values, studies aimed at enhancing bulb production efficiency and increasing the accumulation of pharmaceutically important secondary metabolites are essential.
Eleutherine bulbosa bulbs are composed of a vertically compressed stem with apical and axillary buds encircled by multiple layers of overlapping modified leaves (enlarged leaf bases) that resemble onions and serve as the main organs for storage (Borges et al., 2020; Tribble et al., 2021). Bulbs can store carbohydrates and other nutrients due to thickening development, which requires cell expansion following proliferation at different lateral meristems (Plunkert et al., 2024). Bulb initiation and growth are regulated by exogenous and environmental factors such as phytohormones. Phytohormones regulate plant growth and are thought to have a crucial role in bulb formation and bulb enlargement (Atif et al., 2020). Gibberellins are among the most important (Shu et al., 2024). A phytohormone that promotes growth, gibberellins increase the quantity of cloves per bulb and boost their production efficiency by promoting cell elongation, leaf expansion, axillary meristem initiation, and lateral bud creation (Liu et al., 2020; Khokhar, 2023; Ritonga et al., 2023). Furthermore, GA3 can promote the growth of lateral buds and enhance tuber count (Liu et al., 2019). Therefore, applying GA3 at appropriate concentrations to affect bulblet formation and number, as well as the storage compounds in the bulb, is the main objective of this study.
2. MATERIALS AND METHODS
2.1. Materials
■ E. bulbosa mother bulbs from the National Institute of Medical Materials (Figure 2A and B)
■ The 6-, 12-, and 18-week-old E. bulbosa were grown in the experimental garden of the Department of Plant Physiology, University of Sciences, Vietnam National University, Ho Chi Minh City (Figures 1, 2C, D, and E)

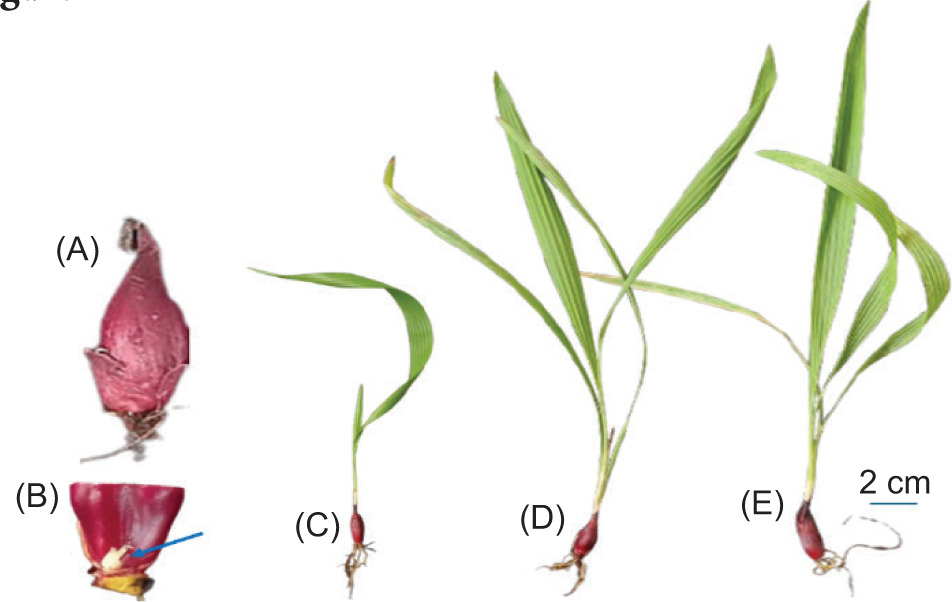
3. METHODS
3.1. Study on the effect of GA3 on E. bulbosa bulblet formation from mother bulbs
E. bulbosa mother bulbs were treated with GA3 by soaking in GA3 solution at different concentrations (from 20 to 50 ppm) (0.5 cm from the base) for a week, then placed in pots with a volume of (20 cm × 21 cm × 20 cm) containing organic soil and coconut fiber mixed in a 1:1 ratio. After 12 weeks, the following parameters were determined: number of bulblets, diameter, fresh weight, dry weight, bulb length, respiration intensity, total sugar content, flavonoid, and polyphenol content (PC).
3.2. Study on the effect of GA3 after 6 weeks of treatments at the 6-week-old plant stage of E. bulbosa
E. bulbosa mother bulbs had the outer dry leaf scales removed, the leaf top and old roots cut off, and were soaked in water (0.5 cm from the base) for a week. After a week, the mother bulbs developed about 2 leaves and 7–10 roots, which were placed in plastic cups with a volume of (95 mm × 58 mm × 125 mm), containing organic soil and coconut fiber mixed in a 1:1 ratio. After 6 weeks of planting, the seedlings from the original mother bulbs were transferred to pots (20 cm × 21 cm × 20 cm) to adapt for 3 days and treated with GA3 at different concentrations (from 20 to 50 ppm). The parameters, including number of bulblets, bulb diameter, fresh weight, dry weight, and length, total sugar content, respiration intensity, flavonoid, and PC in bulb on week 12th, were determined.
3.3. Study on the effect of GA3 on the growth stages of E. bulbosa
E. bulbosa mother bulbs were treated with GA3 by soaking in GA3 30 ppm for a week, then placed in pots with a volume of (20 cm × 21 cm × 20 cm) containing organic soil and coconut fiber mixed in a 1:1 ratio. A 40 ppm GA3 treatment was continued at weeks 6, 12, and 18. The experiment consisted of six treatments (Table 1). The parameters for fresh weight, dry weight, number of bulbs, bulb yield per pot, flavonoid, and PC were determined at the 24th week of treatment.
Table 1
Processing method effect of GA3 on the growth stages of E. bulbosa.
3.4. Analysis of physiological and biochemical parameters
■ Determination of the diameter and length of the bulb: The diameter of the bulb is measured with a ruler, clamped at the largest area of edema, and the length of the bulb is measured with a ruler in millimeters, from the position of the root to the position where the leaf sheath separates.
■ Determination of fresh and dry weight of bulbs: E. bulbosa bulbs were weighed using an analytical balance with an error of 0.01 mg of OHAUS (USA). Then, dried at 120 oC for 1 hour, then at 80 oC for about 72 hours until the weight remained unchanged to determine the dry weight (Sade et al., 2015).
■ Determination of total sugar content of bulbs: Total sugar content in E. bulbosa bulb were determined using the phenol-sulfuric acid method according to Combs et al. (1987) and González-Vázquez (2022). Absorbance was measured at a wavelength of 490 nm using a spectrophotometer, compared to the sucrose standard curve to obtain the total sugar content.
■ Measurement of respiratory intensity: E. bulbosa bulb during treatment was determined using a Leaf Lab 2 oxygen electrode gas exchanger (Hansatech, UK) with an improved measuring chamber with a height of 6 cm. Oxygen exchange rate was calculated based on the amount of oxygen absorbed per gram of fresh weight per minute μmol O2 g–1 min–1.
■ Determination of total phenolic and flavonoid content Polyphenol and flavonoid of bulb dried powder were extracted in 60 % ethanol at a ratio of 1:35 (w/v) (at pH 3.0) for 1 h at 50 oC, then the extract was filtered and allowed to dry. PC was determined spectrophotometrically using the Folin–Ciocalteu method described by Shi et al. (2019), which used a color reaction with Folin-Ciocalteu reagent and 20 % Na2CO3 with gallic acid as the standard, then measured OD at 765 nm. Aluminum chloride spectrophotometry was used to assess total flavonoid content (TFC), as described by Atanassova et al. (2011), which involved a color reaction with 5 % NaNO2 and 10 % AlCl3, with OD measured at 510 nm. Rutin was used as a standard. Polyphenol and flavonoid content in bulb were expressed as mg/g dry weight.
■ Experimental design and statistical analysis
The plants are placed in the experimental garden under light intensity of 10,000–25,000 lux, temperature of 33 ± 5 °C, and humidity of 60 – 65 %. Ten replicates of three plants each were used in the fully randomized design of the studies. Plants were fertilized with NPK 16:16:16 (starting at week 4, once every 4 weeks). Watering was performed once daily at 4 p.m., with a uniform spray until the soil and leaves were wet.
SPSS (Statistical Package for the Social Sciences) 20.0 was used to analyze the data. Statistical differences were significant at the 95 % level, and results were expressed as means ± standard deviations, with accompanying text samples.
4. RESULTS
4.1. Effect of GA3 on growth and bulblet formation of E. bulbosa
After 12 weeks of treatment, in the 30 ppm GA3 treatment, the number of tubers increased rapidly to reach 6–7 tubers, much higher than the control (2–3 bulbs) and other treatments (2–3 or 5–6 bulbs) (Figure 3A–E). The number of bulbs, bulb fresh weight, and dry weight increased sharply at 30 ppm GA3 treatment, then declined at 40 ppm and dropped markedly at 50 ppm. Bulb diameter was reduced in all treatments compared to the control, and the sharpest drop was observed in the 50 ppm GA3 treatment (Table 2 and Figure 3 A1E1). Bulb length also increased sharply at 30 ppm and 40 ppm GA3 treatment but decreased at 50 ppm GA3 treatment (Table 2).
Figure 3
Eleutherine bulbosa after 12 weeks of treatments, with (A): Control; (B): GA3 20 ppm; (C): GA3 30 ppm; (D): GA3 40 ppm; (E): GA3 50 ppm and horizontal cutting bulb detail with (A1) Control (A1); (B1): GA3 20 ppm; (C1): GA3 30 ppm; (D1): GA3 40 ppm; (E1): GA3 50 ppm.
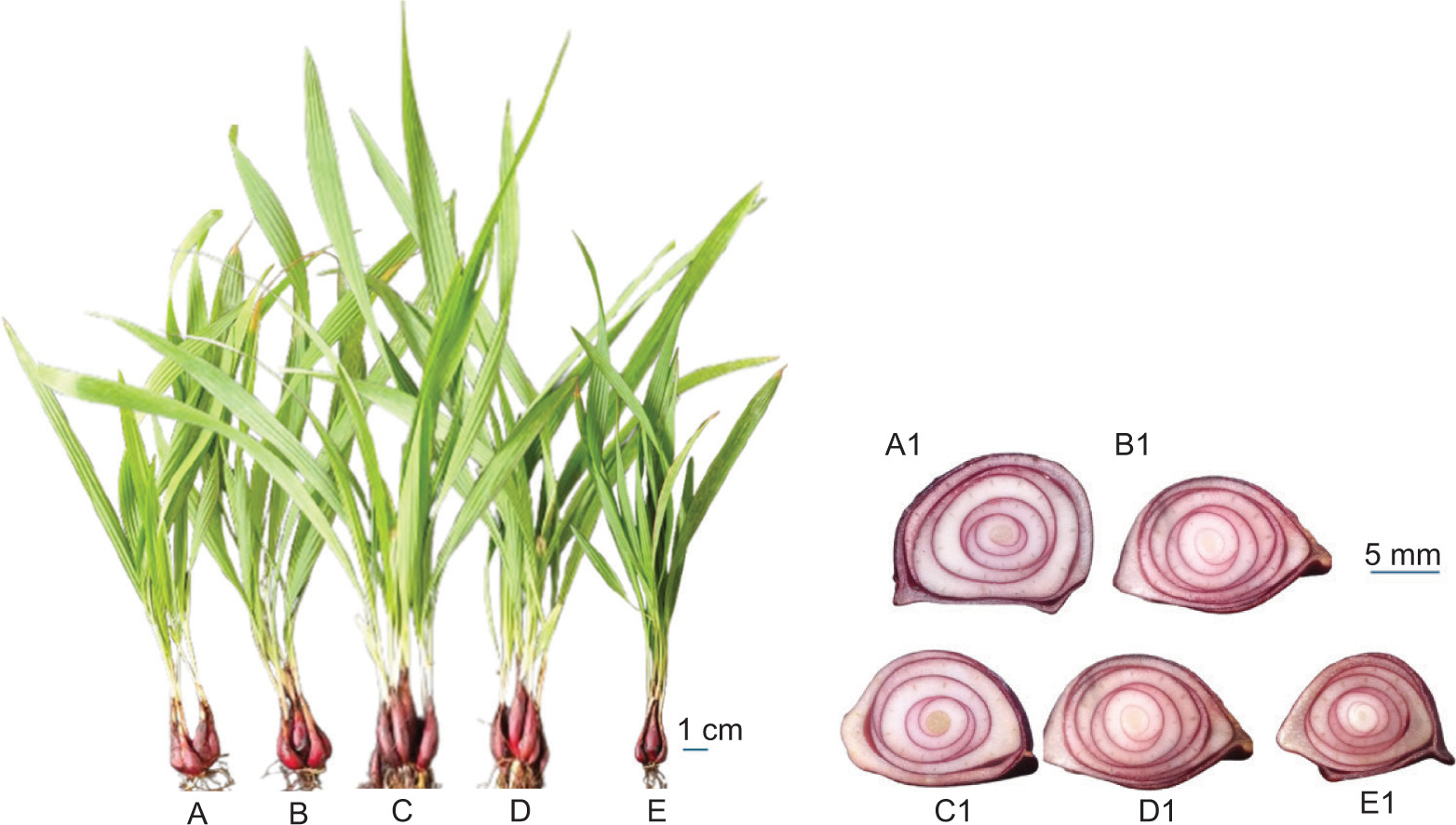
Table 2
Changes in bulb diameter, fresh weight (FW), dry weight (DW), bulb length, and the number of leaf sheaths of E. bulbosa from mother bulbs after 12 weeks of treatments
The sugar and starch content decreased from 20 ppm to 30 ppm after GA3 treatment and then remained constant at 50 ppm treatment. The polyphenol and flavonoid levels began to increase at 30 ppm GA3 treatment and then remained constant at 50 ppm GA3 treatment. The respiration intensity of bulbs also increased sharply at 30 ppm GA3 treatment and then decreased, especially at 50 ppm GA3 treatment (Table 3).
Table 3
Total sugar content (TSC), starch content (StC), flavonoids, polyphenols content, and respiration intensity (RI) of E. bulbosa bulbs after 12 weeks of treatment with GA3.
4.2. Effect of GA3 after 6 weeks of treatments at the 6-week-old plant stage of E. bulbosa
After 6 weeks of treatment at the 6-week-old plant stage, there was no significant difference in bulb count between the control and the treatments (Figure 4A–E), except for the 50 ppm GA3 treatment, which had fewer bulbs. The bulb diameter, fresh weight, and dry weight gradually increased with the GA3 treatment concentration and peaked at the GA3 40 ppm, after which they declined at 50 ppm (Table 3 and Figure 3A1– E1). Similarly, bulb length increased most at 30 and 40 ppm GA3 and decreased sharply at 50 ppm GA3 (Table 3). Sugar and starch contents were reduced in all treatments relative to the control. The starch content plummeted at the 40 ppm GA3 treatment and then remained unchanged at the 50 ppm GA3 treatment, whereas the sugar content plummeted at the 50 ppm GA3 treatment. The polyphenol and flavonoid contents increased sharply at 40 ppm GA3 treatment and then remained constant at 50 ppm GA3 treatment (Table 5).
Figure 4
E. bulbosa after 6 weeks of treatments, with (A1): Control; (B1): GA3 20 ppm; (C1): GA3 30 ppm; (D1): GA3 40 ppm; (E1): GA3 50 ppm and horizontal cutting bulb detail with (A1) Control; (B1): GA3 20 ppm; (C1): GA3 30 ppm; (D1): GA3 40 ppm; (E1): GA3 50 ppm.
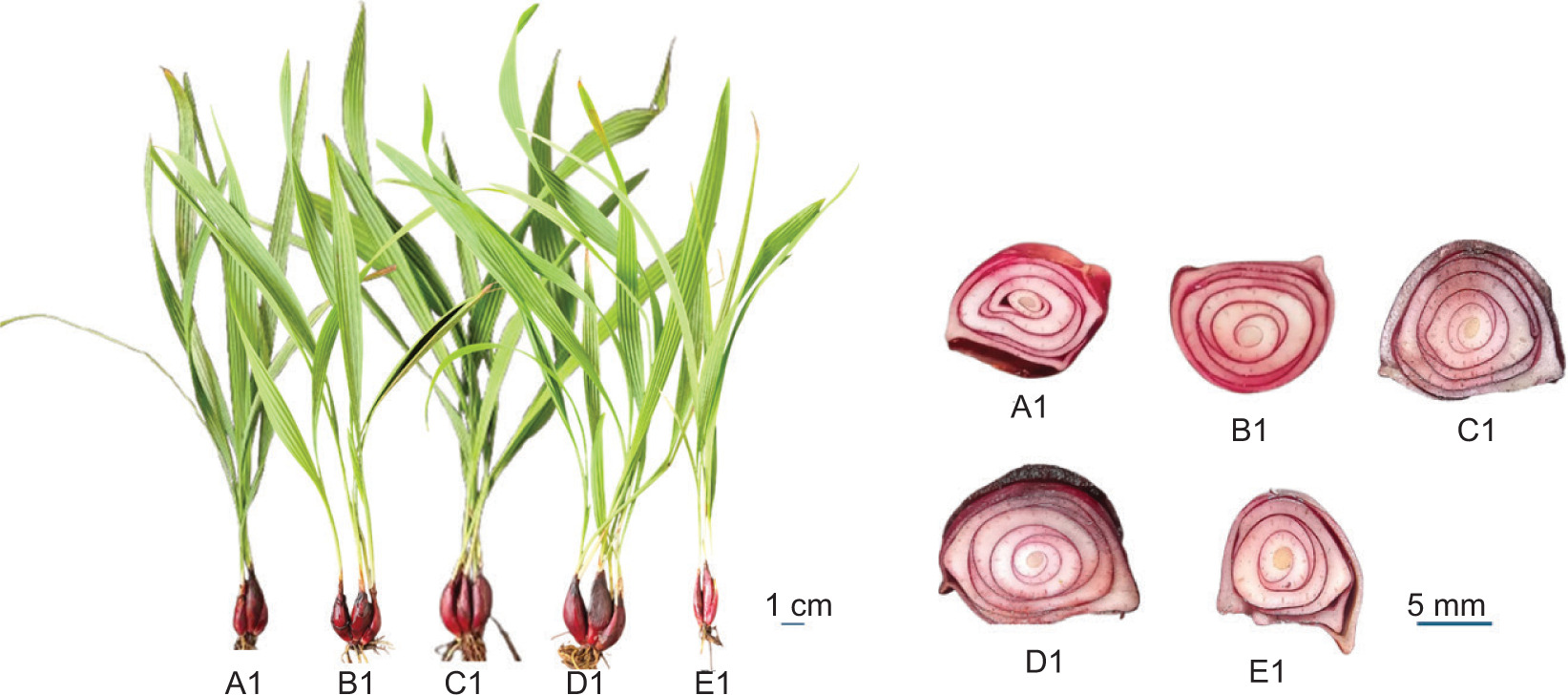
Table 4
Changes in bulb diameter, fresh weight (FW), dry weight (DW), bulb length, and the number of leaf sheaths of E. bulbosa from after 6 weeks of treatments at the 6-week-old plant stage.
Table 5
Total sugar content (TSC), starch content (StC), flavonoids, polyphenols content, and respiration intensity (RI) of E. bulbosa bulbs after 6 weeks of treatment with GA3 at the 6-week-old plant stage.
4.3. Effect of GA3 combined treatments on the growth stages of E. bulbosa
The fresh and dried weight of the bulbs increased in all treatments compared to the control. Between the 2 control experiments, Control 2, which was treated with 30 ppm GA3 at week 0 (mother bulb), had a significantly higher fresh and dry weight. There was no difference in fresh and dry weight between GA3 treatments at weeks 6, 12, and 18. When combined treatments were applied at three stages, these indicators reached their highest values. Likewise, flavonoid and PCs and tuber yield per pot also showed the greatest increases under the combined-stage treatment. The polyphenol content (PC) in Control 2 increased compared to Control 1, but flavonoid content did not differ. Comparing single-stage applications showed that GA3 at week 12 and week 18 did not differ, and both increased markedly versus week 6, while PC at week 18 increased sharply compared with the other 2 weeks. However, bulb yield per pot did not differ among single-stage treatments (Table 6).
Table 6
Changes in bulb fresh weight (FW), dry weight (DW), flavonoid content, polyphenol content, and bulb yield per plant of E. bulbosa after 12 weeks in treatments GA3 at week 12 (the 24-week-old plant).
5. DISCUSSION
5.1. Effect of GA3 on growth and bulblet formation
Eleutherine bulbosa bud derived from mother bulbs after 12 weeks of GA3 treatment showed that the number of bulb-lets, bulb length, and both fresh and dry weight gradually increased at GA3 concentrations of 20–30 ppm, with a significant improvement recorded at 30 ppm compared with the control. In contrast, a higher level (50 ppm) caused a decrease in these parameters (Table 2). This suggests that GA3, like many other plant growth regulators, has a concentration-dependent dual effect. At moderate levels, it promotes axillary meristem initiation and stimulates cell elongation (Liu et al., 2020), thereby enhancing bulblet formation and elongation in E. bulbusa, which contributes to increased fresh weight. However, when the hormone is applied at excessive concentrations, growth parameters are suppressed. These results are similar to reports on garlic and lily that GA3 at appropriate levels enhanced bulblet production, but excessive application reduced yield (Khokhar, 2023; Anisah et al., 2023).
Furthermore, during the early growth stage of E. bulbosa, exogenous GA3 application may impact both bulblet initiation and longitudinal cell expansion within the bulb. This results in an increase in bulblet number and bulb length but is often accompanied by a reduction in bulb diameter, as studied in Tulipa saxatilis (Sari, 2024). This may be explained by the fact that gibberellins regulate growth mainly by stimulating structural proteins and enzymes to promote cell elongation (Kou et al., 2021), rather than enhancing radial expansion.
The effect of GA3 on bulb information and growth in E. bulbosa also depends on the bulbs’ growth stage. At the early stage of growth 0 week (mother bulb stage), the optimal concentration for improving growth goals is GA3 30 ppm, especially increasing the number of bulbs while at the 6 week stage, GA3 at 40 ppm significantly increases fresh weight, dry weight, and length, but does not increase the number of bulbs, and at 50 ppm all parameters decrease (Figures 3 and 4; Table 4). This indicates that in later stages of growth, GA3 favors biomass accumulation rather than only bulblet initiation. These results correspond to the role of GA in promoting cell expansion in preparation for dry matter build-up in the storage organs (Liao et al., 2025).
5.2. Effect of GA3 on secondary compounds
GA3 treatments increase respiration and reduce sugar and starch levels both early from mother bulbs and at 6 weeks (Table 5). This may be because exogenous gibberellin inhibits the activity of sucrosesynthase (SS) and soluble starch synthase (SSS), thereby reducing the sucrose and starch content in the tuber (Li et al., 2021) and increasing the activity of the enzyme responsible for starch hydrolysis, similar to the study by Siega et al. (2025). Moreover, when sugar and starch levels decreased, flavonoid levels in both stages increased, slightly increasing in the early stages, at 30 ppm GA3, sharply increasing in the 6-week stage, at 40–50 ppm GA3, suggesting that carbon reserves were mobilized for new bulblet growth in the early stages and promoted rapid consumption of carbohydrate reserves to provide energy and substrates for organ initiation (Shu et al., 2024), but at a later stage, the metamorphosis shifts towards the biosynthesis of secondary compounds, increasing the content of flavonoids and polyphenols. The effect of GA on flavonoids is due to gibberellin activating phenylalanine aminotransferase activity and promoting the phenylpropanoid metabolism pathway, thereby increasing TFC. Similar to many crops, GA3 promotes the synthesis and accumulation of total phenolic compounds and specific flavonoids (Xu et al., 2025). This is in contrast with some reports that gibberellin signaling suppresses flavonoid biosynthesis in Medicago truncatula or apple (Sun et al., 2021), showing that the effect of GA is species- and stage-dependent.
In the repeated treatment experiments, the combination of 30 ppm GA3 at planting and 40 ppm at later stages (6–18 weeks) resulted in the highest bulb fresh weight, dry weight, bulblet yield, and flavonoid and PCs. This indicates that multiple applications at different developmental stages provide cumulative effects, optimizing both propagation and medicinal quality of bulbs. This once again confirms that the timing of gibberellin application is very important for bulblet bulbosa initiation and enlargement in E. bulbosa, similar to the results of the study in lily (Anisah et al., 2023). Thus, GA3 not only regulates bulblet initiation and biomass accumulation but also promotes the accumulation of flavonoid and polyphenol through carbon mobilization, enhanced respiration, and activation of the phenylpropanoid pathway.
6. CONLUSION
At the mother bulb stage, GA3 at 30 ppm was optimal for increasing bulblet number, fresh and dry weight, bulb length, and respiration intensity. At the 6-week stage, GA3 at 40 ppm enhanced biomass accumulation and secondary metabolite levels, whereas 50 ppm inhibited growth.
Flavonoid and polyphenol contents 9PCs) increased with GA3, reaching the highest levels at 40–50 ppm, depending on growth stage.
Sequential treatments combining 30 ppm at planting with 40 ppm at later stages yielded the greatest improvements in bulb yield and in flavonoid and polyphenol accumulation.
ORCID
Tran Thi Thanh Hien 0009-0002-0213-3440
Vang Thi My Tu 0009-0006-6552-8609
Nguyen Kieu Uyen Vi 0009-0008-2043-1824
Vo Thi Bach Mai 0009-0006-6508-0234
ACKNOWLEDGEMENTS
This research was supported by the Plant Physiology Laboratory, Department of Plant Physiology, Faculty of Biology-Biotechnology, University of Sciences, Vietnam National University in Ho Chi Minh City (VNU-HCM).